https://www.bmj.com/content/2/5040/337
Jo 1957 oli todettu , että idiopaattisen steatorrhean takana voi piillä gluteeni-intoleranssi. Jatkuva steatorrhea aiheuttaa ajanmitatan ADEK-vitamiinien vajautta.
Articles
Effect of a Gluten-free Diet in Idiopathic Steatorrhoea
Ihmisille viljellään sokerimaissia. Löysin tiedon rehumaissista, jota viljellään nautakarjalle. https://yle.fi/uutiset/3-10991952
Artikklein lopussa mainitaan, että ihminenkin voi syödä tätä laatua maissia. olisi mmukava saada vertailu näiden maissilaatujen soveltuvuudesta esim keliakia-diabeteskompbinaatiota poteville. Ja sitäpaitsi keliakiravinto on muutenkin liian glykemistä, joten olisi hyvä löytää vähemmän glykemisiä elintarvikelaatuja.
Celiac
disease (CD) is associated with intestinal lymphoma and other forms of
cancer, especially adenocarcinoma of the small intestine, of the
pharynx, and of the esophagus. Enteropathy-associatedT-cell lymphoma
(EATL) is a rare form of high-grade, T-cell non-Hodgkin lymphoma (NHL)
of the upper small intestine that is specifically associated with CD.
This NHL subtype arises in patients with either previously or
concomitantly diagnosed CD. In a subgroup of patients, there is
progressive deterioration of a refractory form of CD. EATL derives from a
clonal proliferation of intraepithelial lymphocyte s(IEL) and is often
disseminated at diagnosis. Extraintestinal presentations are not
uncommon in the liver/spleen, thyroid, skin, nasal sinus, and brain. The
outlook of EATL is poor. Recent studies indicated that
(1) CD is
associated with a significantly increased risk for NHL, especially of
the T-cell type and primarily localized in the gut (EATL);
(2) the
CD-lymphoma association is less common than previously thought, with a
relative risk close to 3;
(3) CD screening is not required in patients
with NHL of any primary site at the onset, unless suggested by specific
findings (T-cell origin and/or primary gut localization). The risk of
NHL associated with clinically milder (or silent) forms could be lower
than in typical cases of CD. Several follow-up studies suggest that the
GFD protects from cancer development, especially if started during the
first years of life. Strict adherence to the GFD seems to be the only
possibility of preventing a subset of rare but very aggressive forms of
cancer.
References
Fairley, N.H. and Mackie, F.P. The clinical and biochemical syndrome in lymphoma and allied diseases involving the mesenteric lymph glands. BMJ. 1937;
1: 375–380
O’Farrelly, C., Feighery, C., O’Brian, D.S., Stevens, F., Connolly, C.E., McCarthy, C. et al. Humoral response to wheat protein in patients with coeliac disease and enteropathy associated T cell lymphoma. Br Med J. 1986;
293: 908–910
| PubMed.. patients with enteropathy associated T cell lymphoma do not display a
humoral immune response to wheat protein (alpha gliadin), rarely respond
to a gluten free diet, and are often men. Patients with uncomplicated
coeliac disease usually have raised levels of alpha gliadin antibody,
always respond to a gluten free diet, and are frequently women. These
findings suggest the presence of two separate forms of enteropathy: one
is benign and sensitive to wheat protein whereas the other runs a
malignant course.
Fasano, A. and Catassi, C. Current approaches to diagnosis and treatment of celiac disease (an evolving spectrum) . Gastroenterology. 2001;
120: 636–651
Schweizer,
J., Oren, A., Mearin, M.L., and Working Group for Celiac Disease and
Malignancy of the European Society for Pediatric Gastroenterology,
Hepatology and Nutrition. J Pediatr Gastroenterol Nutr. 2001;
33: 97–100
Howdle, P.D., Jalal, P.K., Holmes, G.K.T., and Houlston, R.S. Primary small-bowel malignancy in the UK and its association with coeliac disease. Q J Med. 2003;
96: 345–353
Egan, L.J., Walsh, S.V., Stevens, F.M., Connolly, C.G., Egan, E.L., and McCarthy, C. Coeliac-associated lymphoma. A single institution experience of 30 cases in the combination chemotherapy era. J Clin Gastroenterol. 1995;
21: 123–129
Patey, N., Cellier, C., Jabri, B., Delabesse, E., Verkarre, V., Roche, B. et al. Distinction between coeliac disease and refractory sprue (a simple immunohistochemical method) . Histopathology. 2000;
37: 70–77
We recently showed that refractory sprue
is distinct from coeliac disease, the former being characterized by
abnormal intraepithelial T-lymphocytes expressing a cytoplasmic CD3
chain (CD3c), lacking CD3 and CD8 surface expression, and showing
TCRgamma gene rearrangements. To take advantage of the abnormal
phenotype of CD3c + CD8 - intraepithelial lymphocytes (IEL) in
refractory sprue we developed a simple method to distinguish coeliac
disease from refractory sprue.Comparative
immunohistochemical studies using anti-CD3 and anti-CD8 antibodies were
applied on paraffin-embedded and frozen biopsy specimens in refractory
sprue (n = 6), coeliac disease (n = 10), healthy controls (n = 5) and
suspected refractory sprue (n = 6). Comparable results were obtained on
fixed and frozen biopsy specimens. In four of the six patients with
suspected refractory sprue, abnormal CD3c + CD8 - IEL and TCRgamma gene
rearrangements were found, as in refractory sprue; the remaining two
patients had normal (CD3 + CD8 +) IEL and no TCRgamma gene
rearrangements. Both patients had coeliac disease, as one failed to
comply with a gluten-free diet, while the other was a slow responder.This
simplified immunostaining method using anti-CD3 and anti-CD8 antibodies
on paraffin sections can distinguish active coeliac disease from
refractory sprue and should prove useful in clinical practice.
Harris, O.D., Cooke, W.T., Thompson, H., and Waterhouse, J.A. Malignancy in adult coeliac disease and idiopathic steatorrhoea. Am J Med. 1967;
42: 899–912
| Abstract From a review of 202 patients with adult coeliac disease (ACD) or
idiopathic steatorrhoea (IS) evidence is given that both of these
disorders can be complicated by malignancy, either lymphoma or carcinoma
of the gastrointestinal tract (especially of the oesophagus). The
incidence is highly significant in men, and significant in women. The
mean duration of symptoms of coeliac disease prior to the diagnosis of
lymphoma was 21.2 years, and of carcinoma of the gastrointestinal tract
38.5 years. A gluten-free diet appeared to decrease the risk of
malignant complication.
Askling, J., Linet, M., Gridley, G., Halstensen, T.S., Ekstrom, K., and Ekbom, A. Cancer incidence in a population-based cohort of individuals hospitalised with celiac disease or dermatitis herpetiformis. Gastroenterology. 2002;
123: 1428–1435
Harris, N.L., Jaffe, E.S., Stein, H., Banks, P.M., Chan, J.K., Cleary, M.L. et al. A revised European-American classification of lymphoid neoplasms (a proposal from the International Lymphoma Study Group) . Blood. 1994;
84: 1361–1392
O’Connor, T.M., Cronin, C.C., Loane, J.F., O’Meara, N.M., Firth, R.G., Sanan, F. et al. Type 1 diabetes mellitus, celiac disease, and lymphoma (a report of four cases) . Diabet Med. 1999;
16: 614–617
Bleiberg, H., Duchateau, J., N’Koua M’Bon, J.B., Gerard, B., Bron, D., Debusser, L. et al. Increased incidence of lymphomas and carcinomas in patients with celiac disease. Eur J Cancer. 1998;
34: 592–593
West, J., Logan, R.F.A., Hill, P.G., Lloyd, A., Lewis, S., Hubbard, R. et al. Seroprevalence, correlates, and characteristics of undetected coeliac disease in England. Gut. 2003;
52: 960–965
Obermann, E.C., Diss, T.C., Hamoudi, R.A., Munson, P., Wilkins, B.S., Camozzi, M.L. et al. Loss of heterozygosity at chromosome 9p21 is a frequent finding in enteropathy-type T-cell lymphoma. J Pathol. 2004;
202: 252–262
Peters, U., Askling, J., Gridley, G., Ekbom, A., and Linet, M. Causes of death in patients with celiac disease in a population-based Swedish cohort. Arch Intern Med. 2003;
163: 1566–1572
Corrao, G., Corazza, G.R., Bagnardi, V., Brusco, G., Ciacci, C., Cottone, M. et al. Mortality in patients with coeliac disease and their relatives (a cohort study) . Lancet. 2001;
358: 356–361
Catassi, C., Fabiani, E., Corrao, G., Barbato, M., De Renzo, A., Carella, A.M. et al. Risk of non-Hodgkin lymphoma in celiac disease. JAMA. 2002;
287: 1413–1419
Mearin, M.L. Results
of the European multicenter study on celiac disease and non-Hodgkin
lymphoma (Proceedings of the Xth International Symposium on celiac
disease) . in: John Libbey Eurotext,
Montrouge, France; 2003: 225–227
Farré, C., Domingo-Domenech, E., Font, R., Marques, T., De Sevilla, A.F., Alvaro, T. et al. Celiac disease and lymphoma risk (a multicentre case-control study in Spain) . Dig Dis Sci. 2004;
49: 408–412
1.Clinic and Polyclinic for Internal Medicine II, Klinikum Rechts der IsarTechnical University of Munich MunichGermany 2.Chair of ImmunologyJagiellonian University Medical CollegeKrakówPoland 3.Department of Genetics and DevelopmentColumbia University Medical CenterNew YorkUSA 4.Department of MedicineColumbia University Medical CenterNew YorkUSA 5.Department of UrologyColumbia University Medical CenterNew YorkUSA 6.Herbert Irving Comprehensive Cancer CenterColumbia University Medical CenterNew YorkUSA
Open Access
ReviewFirst Online:
Tiivistelmän suomennosta ja muutamia suomennoksia artikkelin alkuosasta.
. Imettäväiselimistössä on epiteelisolut kaikkein aktiivimmin sykliään tekeviä soluja ja siit johtuen ne ovat altiitta pahanlaatuiselle muuntumiselle. Jo organogeneesin aikana sidekudos eli mesenkyymi tarjoaa opastavia signaaleitaan epiteelille.Aikuiselimistössä uskotaan mesenkyymin tarjoavan ratkaisevia säätelysignaaleita epiteelisolujen ylläpitoon ja uudistumiseen. Tässä artikkelissa pohditaan suoliston myofibroblastien - sileän lihaksen aktiinipositiivisten strooma - eli mesenkyymaalisten alfa-solujen - roolia tärkeänä suoliston kantasolulokeron säätelyosana. Myofibroblastien ja suoliepiteelin välisen kommunikaation ymmärtämisellä on sovellutuksensa, joilla voidaan edistää regeneratiivista lääketiedettä ja parantaa hoitostrategioita tulehduksellisissa suolistorairauksissa, suolistofibroosissa ja kolorektaalisyövässä.
Avainsanoja:
mesenkymaalinen- epiteliaalinen kommunikointi
IBD, inflammatorineneli tulehduksellinen suolistosairaus
myofibroblastit
suolen kantasolulokero
Abstract Epithelial
cells are one of the most actively cycling cells in a mammalian
organism and therefore are prone to malignant transformation. Already
during organogenesis, the connective tissue (mesenchyme) provides
instructive signals for the epithelium. In an adult organism, the
mesenchyme is believed to provide crucial regulatory signals for the
maintenance and regeneration of epithelial cells. Here, we discuss the
role of intestinal myofibroblasts, α-smooth muscle actin-positive
stromal (mesenchymal) cells, as an important regulatory part of the
intestinal stem cell niche. Better understanding of the cross-talk
between myofibroblasts and the epithelium in the intestine has
implications for advances in regenerative medicine, and improved
therapeutic strategies for inflammatory bowel disease, intestinal
fibrosis and colorectal cancer. Keywords: Mesenchymal–epithelial cross-talk Inflammatory bowel disease Myofibroblasts Intestinal stem cells Stem cell niche
JOHDANTO. Kudoksen mikroympäristö
Aikuisen kehossa on epiteliaalisten kantasolujen vastuulla normaalin epiteelikudoksen uudistaminen, esim ihon, hengiysteiden ja mahasuolikanavan pintasolukon regeneroiminen. Epiteelikantasolut ovat jatkuvasti vuorovaikutuksessa paikalliseen ympäristöönsä, joka tunnetaan kantasolujenpesänä, kantasolulokerona ( stem cell niche). tämä ikantasolulokero käsittää extrasellulaarisen matriisin (ECM), liukoisia tekijöitä ja mesenkymaalisia soluja. Mesenkymaalisten solujen joukossa havaitaan esimerkiksi eri tyyppisiä immuunisoluja, endoteelisolujam, neuroneita eli hermosoluja, mesenkymaalisia kantasoluja, fibroblasteja ja myofibroblasteja. Tässä katsauksessa keskitytään mesenkymaalisolujen osuuteen ja erityisesti suoliston myofibroblasteihin (IFM), jotka ovat ratkaisevia komponentteja suoliston kantasolupesässä.
Introduction: Tissue Microenvironment. Epithelial
stem cells are responsible for the normal epithelial tissue
regeneration in an adult organism for example in skin, respiratory tract
and gastrointestinal tract. Epithelial stem cells are constantly
interacting with the local surroundings, known as the stem cell niche,
which is composed of extracellular matrix (ECM), soluble factors and
mesenchymal cells. Among mesenchymal cells, we can distinguish, for
example, different types of immune cells, endothelial cells, neurons,
mesenchymal stem cells, fibroblasts and myofibroblasts. In this review,
we focused on the role of mesenchymal cells, particularly intestinal
myofibroblasts (IMFs), as a crucial component of the intestinal stem
cell niche.
Suolistonkantasolulokero, kantasolujen pesä
Meidän elimistössämme ovat epiteelisolut kaikkein aktiivimmin sykliään
tekeviä soluja ja siitä johtuen altistuneita pahanlaatuiselle
muuntumiselle. Suoliston epiteelin solujen suuren vaihtuvuuden takana on suoliston kantasolulokero, joka sijaitsee suolista krypta-syvennysten pohjalla. Nämä kantasolulokeron solut voidaan erottaa muista soluista, sillä niiden ollessa syklissään aktiiveina niillä on markkerina Lgr5-pintareseptori. Lisäksi uskotaan olevan toinenkin suolistokantasolujen alaryhmä, joka on hiljaiselossa ja sen markkeri on Bmi1 tai muitakin markkereita mainitaan: Hopx, mTERT ja Lrig1. Suoliston epiteeli on heterogeenistä, koska se on koostunut erilaisista epiteelisolutyypeistä kuten enterosyyteistä, enteroendokriinisoluista, gobletin pikarisoluista ja Panethin soluista. Kaikkien näiden solujen alkulähteenä toimivat nämä suolsiton kantasolut (ISC). Aivan viime aikoina on havaittu, että Panethin solut ovat suoliston kantasolulokeron ratkaiseva komponentti ja ne tarjoavat lokeron faktoreita suoliston kantasoluille. Kuitenkaan Panethin solujen vähentäminen ei tehnyt merkitsevää muutosntumista suoliston kryptaan, mikä viittaisi siihen , että mesenkymaaliset solut pitävät huolta toimittaen essentiellejä lokerotekijöitä suoliston kantasoluille.
The Intestinal Stem Cell Niche. Intestinal
epithelial cells are one of the most actively cycling cells in our
body, and they are also prone to malignant transformation. High cell
turnover in the intestinal epithelium is fueled by the intestinal stem
cells (ISCs) that are located at the bottom of the intestinal crypt.
ISCs can be distinguished from other cells by the expression of Lgr5
(Barker et al. 2007),
which marks actively cycling ISCs. In addition, it is believed that
there exists another subpopulation of ISCs that are quiescent and are
marked with Bmi1 (Sangiorgi and Capecchi 2008), or other markers such as: Hopx, mTERT and Lrig1 (Barker et al. 2012).
The intestinal epithelium is heterogenous as it is composed of
different epithelial cell types such as enterocytes, enteroendocrine
cells, goblet cells and Paneth cells. A source for all those epithelial
cell types is an ISC. Recently, Paneth cells have been shown to be a
crucial component of the intestinal stem cell niche and provide niche
factors for ISCs (Sato et al. 2011). However, depletion of Paneth cells did not cause significant alterations in the intestinal crypt (Durand et al. 2012) thus suggesting that mesenchymal cells provide essential niche factors for the ISCs.
Monet tutkimukset ovat antaneet näyttöä mesenkymaalisepiteliaalisen "vuoropuhelun" tärkeydestä suoliston kantasolujen pesässä.
Ensinnäkin Fox11+ mesenkymaaliset solut näyttävät säätelevän suoliston kryptan proliferoitumista (2016).
Toiseksi Wnt5a+ mesenkymaalisten solujen on osoitettu stimuloivan akuutin suolistovaurion mallissa epiteelin uudistumista (2012).
Kolmanneksi kemoterapian indusoiman vaurionkin jälkeisessä suolistoepiteelin toipumisessa on osoitettu mesenkymaalisilla soluilla olevan osuutta(2015).
Lisäksi strooman BMP-signaloinnin osallistumisella on tuntuvaa vaikutusta epiteeliin, koska se jaiheuttaa polyyppien kasvua. ( BMP = luun morfogeneettinen proteiini).
On tärkeä huomata mesenkymaalisepiteeliaalisesta "vuoropuhelusta", että se ei ole yksisuuntaista, vaan suoliston epiteeli myös tarjoaa signaaleita lähistroomaansa (2005), siis suunta on silloin epiteliaalismesenkymaalinen. tässä työssään kirjoittajat osoittivat, että suolsitoepiteelin ilmentavien Shh ja Ihh Hedgehog- signaalitien tekijöiden vähentyminen johtui epiteelinalaisten myofibroblastien väärään lokalisoitumiseen. (Kommentti: Hedgehog kuten Notch ja Wnt kuuluvat kantasolun signalointiteihin ja Hedgehog varsinkin vastaa asianmukaisesta kehityksestä kuten sijoittautumisesta oikeaan lokalisaatioon. Esim Sirtuiinien kirjo taas säätelee näitä tekijöitä, joihin vaikuttaa sirtuiinien suorittama ravinnon ja energian saannin sensorointi).
Mesenchymal–Epithelial Cross-Talk in the Intestinal Stem Cell Niche. Many
studies have provided evidence on the importance of the
mesenchymal–epithelial cross-talk in the intestinal stem cell niche
(Table 1). First, Foxl1+ mesenchymal cells were shown to regulate proliferation in the intestinal crypt (Aoki et al. 2016). Second, Wnt5a+ mesenchymal cells were demonstrated to stimulate epithelial regeneration in an acute intestinal damage model (Miyoshi et al. 2012).
Third, a study of Miyoshi et al. suggests that mesenchymal cells can
also play an important role during intestinal epithelial recovery after
chemotherapy-induced damage (Seiler et al. 2015).
In addition, interference with the bone morphogenetic protein signaling
in the stroma has a profound impact on the epithelium as it results in
the growth of polyps (Beppu et al. 2008).
Importantly, mesenchymal–epithelial cross-talk is not unidirectional,
also the intestinal epithelium provides signals to the adjacent stroma
as it was demonstrated by Madison et al. (2005).
In this study, the authors showed that reduction of Sonic (Shh) and
Indian (Ihh) hedgehog, that are expressed in the intestinal epithelium,
results in mislocalization of subepithelial myofibroblasts (Madison et
al. 2005).
Table 1
Examples of the mesenchymal–epithelial cross-talk in the intestine
Description
References
Deletion of the BMP type II receptor in the stroma induces formation of intestinal polyps
Deletion of Foxl1+ mesenchymal cells reduces epithelial cell proliferation in the intestinal stem cell niche. Moreover, Foxl1+ mesenchymal cells are a source of Wnt ligands in the intestinal stem cell niche
Suoliston kantasolupesässä on fenotyypiltään ja funktioltaan toisistaan erotettavissa olevia mesenkyymisolupopulaatioita kuten alfa-SMA(+) ( sileän lihaksen alfa-aktiini(+) myofibroblasteja (IMF) , alfa-SMA(-) mesenkymaalisoluja, esimerkiksi CD34(+) mesenkymaalisoluja ja Fox11(+) mesenkymaalisoluja. Tässä työssään tutkijat keskittyvät alfa-SMA(+) myofibroblasteihin, koska niitä esiintyy aikuisorganismin lisäksi myös varhaisessa suoliston kehityksessä (2011). Tämä viittaa siihen, että alfa-SMA(+)IMF voi (1) säätää suoliston morfogeneesiä, (2) huolehtia avainasemassa olevista kantasolupesän proliferaatio- ja differentiaatiosignaaleista sekä sikiön että aikuisen suoliston epiteelissä. Lisäksi alfa-SMA(+) myofibroblasteilla on tärkeitä sovelluksia syöpätutkimuksessa.
In the intestinal stem cell niche, there are phenotypically and
functionally distinct populations of mesenchymal cells such as:
alpha-smooth muscle actin (α-SMA)+ myofibroblasts (Powell et al. 1999b) and α-SMA− mesenchymal cells, e.g., CD34+ mesenchymal cells (Stzepourginski et al. 2017) and Foxl1+ mesenchymal cells (Aoki et al. 2016). Here, we focused on the α-SMA+
myofibroblasts, because they are present not only in an adult organism,
but also during early intestinal development (Artells et al. 2011). This suggests that α-SMA+
IMFs could: (1) regulate intestinal morphogenesis; (2) provide key
niche signals for proliferation and differentiation of both fetal and
adult intestinal epithelium. Moreover, α-SMA+ myofibroblasts have important implications for cancer research.
Myofibroblastit ovat sukkulamaisia, supistuvaisia soluja, alunperin mesodermistä ja ilmentävät alfa-SMA.Myofibroblastien vastuulla on tuottaa extrasellulaarisen matriisin (ECM) proteiineja , jotka antavat tukirankaa kudokselle ja kasvutekijöitten signaloinnille (2010). Sen ohella myofibroblastit erittävät laajan kirjon kasvutekijöitä, proteaaseja, sytokiinejä ja kemokiinejä (1999). Myofibroblastit osallistuvat imettäväiselimsitössä moniin prosesseihin. Niillä on myös tärkeä osa kehityksen aikana (2005), angiogeneesissä (2012) ja immuunivasteessa (2007, 2003). Lisäksi ne ovat kriittisiä tekijöitä haavan paranemisessa, josas ne vastaavat vaurioalueen supistumisesta ja arven muodostuksesta(2003, 2013). Monissa taudeissa mainitaan myofibroblastien osuudesta kuten maksakirroosissa, munuasifibroosissa ja keuhkofibroosissa ( 2003, 2011,2013) sekä syövässä. Tuumorisolupesässä myofibroblastit ovat kaikkein runsaimpiin ei-maligneihin solutyyppeihin kuuluvia ja edistävät tuumorin progredioitumista (2012, 2006, 2011). Myofibroblastit on tunnistettu mahdollisina kohteina sekä fibroottisissa taudeissa (2007) että syövässä (2004). Lisäksi suoliston myofibroblasteissa ja kryptan epiteelisoluilla ilmenee Tollin-reseptoreiden kaltaisia reseptoreita , mikä viitaa siihen, että niillä on kykyä kommunikoida suolen mikrobiomin tuotteiden kanssa ja niillä on vaikutusta limakalvon immuniteettiin.
Myofibroblasts
Multiple Functions of Myofibroblasts. Myofibroblast
is a spindle-like, contractile cell that has a mesodermal origin and
expresses α-SMA. Myofibroblasts are responsible for the production of
ECM proteins (Frantz et al. 2010),
which provide a scaffold for the tissue and growth factor signaling.
Besides that, myofibroblasts secrete a broad spectrum of growth factors,
proteases, cytokines, and chemokines (Powell et al. 1999a).
Myofibroblasts are involved in many processes in a mammalian organism.
Myofibroblasts play an important role during development (Mitchell 2005), angiogenesis (Mayrand et al. 2012) and immune response (Andoh et al. 2007; Otte et al. 2003).
Moreover, myofibroblasts are critical players during wound healing,
where they are responsible for contractility of an injured area and
formation of a scar (Gabbiani 2003; Klingberg et al. 2013). Myofibroblasts are implicated in many diseases such as liver cirrhosis, renal fibrosis or lung fibrosis (Gabbiani 2003; Klingberg et al. 2013; Meran and Steadman 2011),
and cancer. At the tumor niche, myofibroblasts are one of the most
abundant non-malignant cell type and promote tumor progression (Cirri
and Chiarugi 2012; Orimo and Weinberg 2006; Quante et al. 2011). Myofibroblasts are recognized as potential targets for both fibrotic diseases (Scotton and Chambers 2007) and cancer (Micke and Ostman 2004).
Moreover, IMFs along with crypt epithelial cells express Toll-like
receptors that points to their ability to cross-talk with gut microbiota
products and their impact on mucosal immunity (Brown et al. 2014).
Suoliston epiteelin alaiset myofibroblastit
Suolistossa aivan suoliston epiteelin lhellä sijaitsevia myofibrolasteja tunentaan subepiteelisinä myofibrobalsteina tai perikryptisinä myofibroblasteina. Suoliston krypta muodostuu noin 250 epiteelisolusta ja niissä on 15 Lgr5(+) kantasolua (2013). Joka päivä noin 200 uutta kryptaa kehkeytyy. Ohutsuolessa noin 38 myofibroblstia ja paksusuolessa 124 myofibroblastia muodostavat kotelon, kryptan ympärille ( muodostaen kantasolupesän)(1981). Nuo myofibroblastit ovat alfa-SMA(+), vimentiini(+) ja desmiini(-) soluja ja ne suorittavat hidasta sykliään ja fusoituvat toinen toisiinsa muodostaen synsytiumin (1999) . Tuore tutkimus (2017) viittaa siihen, että miRNA204 ja 211 pystyvät tekemään eron subepiteliaalisten mikroblastien ja alfa-SMA(-)mesenkymaalisten stroomasolujen välillä. Siitä huolimatta sekä mikroRNA:t että hyvin tunnetut mesenkymaaliset stroomasolumarkkerit ( kuten alfa-SMA, vimentiini ja desmiini) ilmentävät solunsisäistä paikallistumista. Täten on kiireellistä tarvetta tunnistaa uusia stroomasolumarkkereita, jotka kuuluvat solupintaproteiinien ryhmään, jotta niitä voisi käyttää hiiren fluoresaatiolla aktivoituvien solujen lajittelussa (FACS)- samoin myös ihmiskudoksen tutkimuksissa, jolloin voitaisiin nopeuttaa stroomasolujen osuuden ymmärtämistä gastrointestinaalisissa kroonisissa taudeissa.
Transplantaatiotutkimukset ovat osoittaneet , että niin hiiren kuin ihmisen suoliston subepiteliaaliset myofibroblastit ovat alkuisin luuytimestä (2002). Sen lisäksi myofibroblastit voivat olla peräisin paikallisista fibrioblasteista ja paikallisista mesenkymaalisista kantasoluista , suoliston retikulaarisista gremlin (+) kantasoluista, fibrosyyteistä ja myös EMT:stä , epiteliaalisesta mesenkymaaliseen siirtymisestä johtuen (2011-2015). Suoliston myofibroblasteja alkaa ilmetä ensimmäisen kerran ihmisellä kehityksen 9. viikon aikana (2011). Mielenkiintoinen havainto on , että myofibroblastien ilmestyminen korreloi suoliston ontelon muodostumiseen (2011), mikä viittaa tämän stroomasolutyypin olevan ehkä ratkaisevassa osassa suoliston epiteelin morfogeneesin aikana.
Subepithelial Myofibroblasts in the Intestine. In
the intestine, those myofibroblasts that are adjacent to the intestinal
epithelium are known as subepithelial myofibroblasts or pericryptal
myofibroblasts. The intestinal crypt is composed of about 250 epithelial
cells, including 15 Lgr5+ stem cells (Clevers 2013).
Each day about 200 new crypts are generated. About 38 myofibroblasts in
the small intestine and 124 myofibroblasts in colon form a niche around
a crypt (Neal and Potten 1981). Those myofibroblasts are α-SMA+, vimentin+ and desmin− cells, and are slowly cycling, and fuse with each other to form syncytia (Powell et al. 1999b). A recent study of Sacchetti et al. (2017) suggests that expression of microRNA-204&211 can distinguish subepithelial myofibroblasts from α-SMA−
mesenchymal stromal cells. Nevertheless, both microRNAs as well as
well-known mesenchymal cell markers, e.g., α-SMA, vimentin and desmin,
exhibit intracellular localization. Hence, there is an urgent need to
identify novel stromal cell markers that belong to the group of cell
surface proteins, so that they could be used for fluorescence-activated
cell sorting (FACS) of the mouse as well as human tissue that will
certainly accelerate progress in understating the contribution of
stromal cells to chronic diseases of the gastrointestinal tract. Transplantation
studies demonstrated that subepithelial myofibroblasts in the intestine
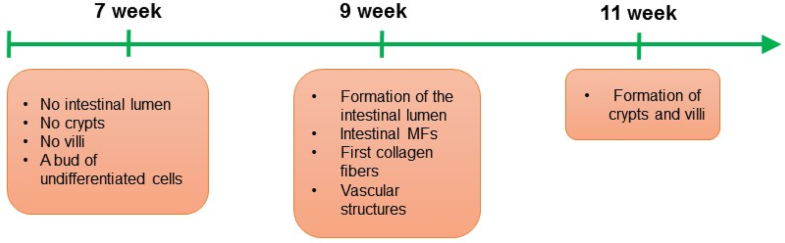
in both mice and human originate from bone marrow (Brittan et al. 2002). Besides that, myofibroblasts can originate from local fibroblasts and local mesenchymal stem cells, gremlin+ intestinal reticular stem cells, fibrocytes, and as result of the epithelial–mesenchymal transition (EMT) (Artells et al. 2011; Micallef et al. 2012; Worthley et al. 2015). IMFs appear for the first time during the 9 weeks of human development (Artells et al. 2011). Excitingly, appearance of myofibroblasts correlates with formation of the intestinal lumen (Artells et al. 2011) (Fig. 1), which implies that this stromal cell type can play a crucial role during the intestinal epithelial morphogenesis.
-
Fig. 1
Organogenesis
of human small intestine and initiation of the myofibroblast
(MF)–epithelium interactions in the intestinal stem cell niche. During
the 7 weeks of small intestine human development, a bud of
undifferentiated cells is observed, at that time point crypts and villi
are not formed yet. During the 9 weeks of small intestine human
development, the intestinal lumen is initiated, and the first intestinal
MFs, vascular structures and collagen fibers are detected. During the
9 weeks of small intestine human development crypts and villi are
present
.Presence of IMFs during early intestinal organogenesis and their
subepithelial localization in the adult intestine suggests that these
mesenchymal cells may provide some crucial niche factors for the ISCs,
and regulate proliferation and differentiation in the intestinal
epithelium. Indeed, in situ hybridization revealed that subepithelial
myofibroblasts can express Wnt ligands such as Wnt2b, Wnt4 and Wnt5b
(Gregorieff et al. 2005),
which strongly suggests that this stromal cell type can regulate Wnt
signaling in the adjacent epithelial cells. Wnt pathway provides
essential signals for ISCs and deregulations in this pathway are
associated with the development of the intestinal cancer (Reya and
Clevers 2005).
However, surprisingly, the study of San Roman et al. has shown that
deletion of porcupine (an enzyme responsible for posttranslational
modifications and Wnt secretion) in Myh11+ cells (that include subepithelial myofibroblasts) has no phenotype in the intestinal crypt (San Roman et al. 2014)
suggesting that there can be other niche cells compensating for the
loss of Wnt secretion in subepithelial myofibroblasts. A possible
explanation is, e.g., the presence of CD34+ mesenchymal cells (Stzepourginski et al. 2017) and Foxl1+ mesenchymal cells (Aoki et al. 2016) (Fig. 2).
These cell types could provide compensatory signals, including Wnt
ligands, for the ISCs in the absence of functional Wnts in the Myh11+ cells. Of note, CD34+ mesenchymal cells described by Stzepourginski et al. (2017) were studied only in ileum and colon. Moreover, Gli1+ subepithelial mesenchymal cells were proposed to be a source of Wnt ligands in the intestinal stem cell niche (Valenta et al. 2016);
however, this stromal cell subpopulation remains uncharacterized. It is
worth to add that besides mesenchymal cells, Paneth cells also can be a
source of Wnt ligands for the ISCs, e.g., Wnt3 (Sato et al. 2011).
Interestingly, depletion of Paneth cells has no phenotype in the
intestinal crypt under homeostatic conditions (Durand et al. 2012).
Altogether, this can suggest a cooperative work of epithelial (such as
Paneth cells) and stromal cells in the intestinal stem cell niche.
Existence of redundant mechanisms to maintain ISCs could protect against
the loss of ISCs, which are necessary to maintain the pool of
enterocytes whose primary function is nutrient absorption.
.
-
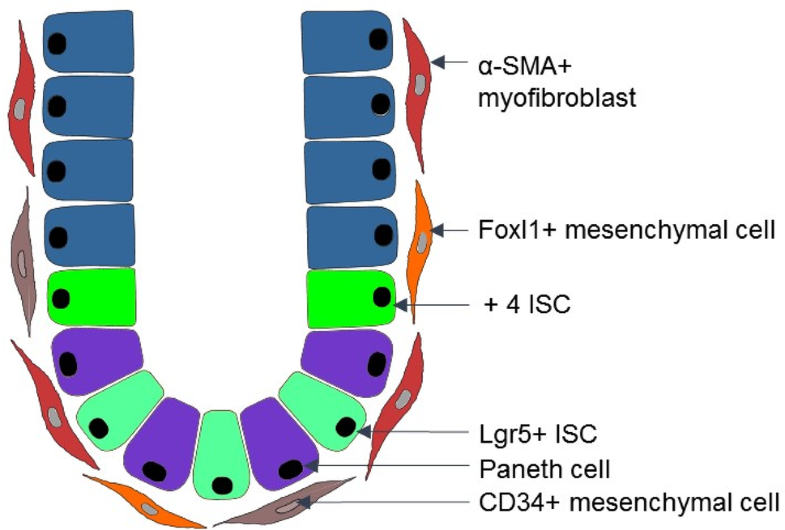
Fig. 2
Scheme
of the mesenchymal niche in the intestine. In the intestinal crypt,
there are at least two subpopulations of intestinal stem cells (ISCs):
Lgr5+ ISCs and + 4 ISCs that are responsible for the high
regeneration capacity of the intestinal epithelium. Crypt cells,
including ISCs, are in close contact with different types of mesenchymal
cells such as: CD34+ mesenchymal cells, Foxl1+ mesenchymal cells and α-SMA+ myofibroblasts
Although, for many years it has been believed
that Wnts are critical regulators of the epithelial self-renewal in the
intestinal crypt, data from in vitro (Glinka et al. 2011) and in vivo studies (Yan et al. 2017)
revealed that in addition to Wnt ligands, R-Spondins also play a
critical role in Wnt pathway. R-Spondins are secreted proteins that are
involved in maintenance of the surface localization of a receptor-bound
Wnt through regulation of the transmembrane E3 ligasesRnf43/Znrf3,
which ultimately results in amplification of the Wnt signal (Farin et
al. 2016). Yan et al. (2017) proposed that not Wnts, but rather R-Spondins may play a dominant role in self-renewal of Lgr5+ intestinal stem cells. Interestingly, R-Spondins are likely produced by stromal cells (Sigal et al. 2017); nonetheless, this requires detailed investigation in the future.
Besides
Wnt ligands and R-Spondins, many other niche signals were shown to
regulate intestinal epithelial cells. Here, among the molecules involved
in the intestinal (myo)fibroblast—intestinal epithelial cell cross-talk
are, e.g., hepatocyte growth factor (HGF) (Goke et al. 1998), prostaglandin E2 (PGE2) (Roulis et al. 2014), and periostin (Kikuchi et al. 2008).
Moreover, IMFs, together with smooth muscle cells, were shown to guide
intestinal epithelial regeneration in a dextran sulfate sodium (DSS)
injury model via mechanism that involves microRNA-143/145 and
insulin-like growth factor binding protein 5 (IGFBP5) (Chivukula et al. 2014).
Furthermore, very recently angiopoietin-like protein 2, that is
expressed in subepithelial myofibroblasts in colon, was demonstrated to
play an important role during regeneration of the intestinal epithelium
in two mouse models of intestinal injury (Horiguchi et al. 2017). Altogether, this suggests that IMFs regulate intestinal epithelial cells via various molecular mechanisms.
IMFs in Disease
Increased number of α-SMA+ myofibroblasts was observed during both intestinal inflammation and intestinal tumor (Andoh et al. 2002; Powell et al. 2005). The role of IMFs during disease was previously thoroughly reviewed, e.g., by Powell et al. (2011), Roulis and Flavell (2016) and Koliaraki et al. (2017).
Importantly, during intestinal inflammation and cancer, not only the
number of IMFs is altered, but also changes in gene expression profiling
and proteome profiling were detected in IMFs. For example, increased
expression of inflammatory mediators such as interleukin 6, osteopontin,
CXCL2 and CCL20 was found in carcinoma-associated fibroblasts (CAFs)
derived from azoxymethane/dextran sodium sulfate (AOM/DSS) mice, an in
vivo model of colitis-associated cancer, when compared to normal
myofibroblasts (Torres et al. 2013).
In addition, using the same research model, it was shown that tumor
progression locus 2, a kinase that is expressed in IMFs, protects
against colitis-associated cancer by regulating production of HGF
(Koliaraki et al. 2012).
Similarly, epimorphin, a mesenchymal protein, was shown to exhibit a
protective role against colitis-associated cancer in AOM/DSS mouse model
(Shaker et al. 2010).
The potential limitations of the studies above are that: (1) AOM/DSS
mouse model might not recapitulate the genetic landscape of human
colitis-associated colorectal cancer, and (2) the differences between
mouse and human immune system. A potential solution here is the
application of human-derived organoid models and mouse models with
humanized immune system to understand better the epithelial–stroma
interactions in colitis-associated cancer.
Immunohistochemistry and gene expression data provided evidence that
myofibroblasts could serve as a prognostic factor in colorectal cancer
(Isella et al. 2015; Tsujino et al. 2007).
It is worth to mention that in case of the global gene expression
analyses of colorectal tumor tissue, myofibroblasts can be a source of
“pseudo-EMT signals” (Calon et al. 2015),
which should be taken into consideration when analyzing any gene
expression data obtained from the whole tumor tissue. Additionally,
stromal microRNA-21 was shown to have prognostic value in colorectal
cancer (Nielsen et al. 2011). Excitingly, such stromal microRNA-21 can be associated with exosomes (Bhome et al. 2017),
a type of extracellular vesicles that are produced by mammalian cells
for the intercellular communication. Interestingly, CAF-derived exosomal
microRNA-21 was shown to have an impact on colorectal cancer cell
proliferation, resistance to chemotherapy and formation of liver
metastases (Bhome et al. 2017).
Besides that, mechanistically, myofibroblasts isolated from colon
cancer tissue were shown to promote tumor cell invasion via mechanism
that involves tenascin-C, scatter factor/HGF, RhoA and Rac (De Wever et
al. 2004). Moreover, a study of Vermeulen et al. (2010) suggests that myofibroblasts could contribute to the “β-catenin paradox” (mosaic pattern of β-catenin nuclear localization) observed in colorectal cancer cells.
Inflammatory bowel disease (IBD) is characterized by epithelial injury
and intestinal inflammation. IBD is a group of diseases that include
ulcerative colitis and Crohn’s disease. One of the key cytokines that is
involved in the pathogenesis of IBD is IL-33, which belongs to the IL-1
superfamily of cytokines; IL-33 is responsible for immune cell
infiltration and Th2 responses (Miller 2011; Neurath 2014). Interestingly, the study of Sponheim et al. (2010)
suggests that pericryptal myofibroblasts are a source of IL-33 in
patients with ulcerative colitis, which highlights an important role of
this cell type in the pathogenesis of ulcerative colitis and warrants
for further studies on the role of pericryptal myofibroblasts in IBD.
Moreover, the study of Messina et al. (2017) suggests that colonic CD146+ cells, that were shown to have features of IMFs (Signore et al. 2012),
exhibit increased expression of HLA-DR, a major histocompatibility
complex class II antigen. However, this requires more investigation. The
findings should be confirmed using larger number of samples and
functional studies should be performed. Additionally, it was shown that
human IBD IMFs exhibit differential expression of distinct transforming
growth factor β isoforms (McKaig et al. 2002).
To summarize, IMFs are an important component of the stromal niche
during IBD and intestinal cancer. Better understanding of the role of
IMFs during pathogenesis of IBD and intestinal tumor can potentially
lead to identification of new therapeutic targets for those diseases.
Summary and Future Directions
To
summarize, emerging data highlight the importance of
mesenchymal–epithelial cross-talk in the intestine during homeostasis,
regeneration after an injury and chronic diseases. Here, we particularly
focused on subepithelial myofibroblasts that surround the intestinal
crypt. Many studies pointed out the important role of the subepithelial
myofibroblasts in regulation of intestinal epithelial proliferation via
different molecular mechanisms that involve, e.g., HGF, PGE2,
periostin, microRNA-143/145 and IGFBP5. Still, many questions remain to
be answered. For example, it would be interesting to decipher whether
subepithelial myofibroblasts can activate quiescent ISCs and if
migration of crypt cells along the crypt–villus is regulated
autonomously or rather by subepithelial myofibroblasts?. In a mammalian
organism, there are multiple mechanisms responsible for the maintenance
of adult stem cells. One example is asymmetric organelle segregation
during cell division (Ouellet and Barral 2012). The study of Katajisto et al. (2015)
demonstrated that young mitochondria are preferentially distributed to
stemlike cells during mitosis of mammary epithelial cells. It would be
exciting to unpuzzle whether subepithelial myofibroblasts could regulate
segregation of mitochondria in the neighboring ISCs. Given the stromal
cell heterogeneity in the intestinal stem cell niche, it would be also
interesting to study the relationship of subepithelial myofibroblasts
with other types of mesenchymal cells such as CD34+ mesenchymal cells and Foxl1+ mesenchymal cells. Moreover, differentiation status of CD34+ and Foxl1+
mesenchymal cells remains unclear: can these cell types act as
progenitor cells for the myofibroblast syncytium?. It is also unknown
whether CD34+ mesenchymal cells are the same cells as Foxl1+ mesenchymal cells.
Aberrant
niche signaling was detected in various human diseases, including
colorectal cancer and IBD. IMFs were identified as one of key components
of the stromal niche in both colorectal cancer and IBD, where IMFs were
suggested, e.g., to be a source of inflammatory mediators. Future
studies should provide more input into the precise role of subepithelial
myofibroblasts in the regulation of immune response in IBD. It would be
also interesting to study whether subepithelial myofibroblasts can
provide signals promoting self-renewal of colon cancer stem cells. And,
as niche factors are especially important during epithelial homeostasis
and very early stages of intestinal tumor growth (Fujii et al. 2016),
it would also be intriguing to investigate whether
myofibroblast-derived niche factors can promote tumor initiation process
in the intestinal epithelium. Overall, increasing the knowledge on the
myofibroblasts–intestinal epithelium cross-talk in the intestinal stem
cell niche during homeostasis and disease can lead to identification of
novel therapeutic targets, e.g., for colon cancer and IBD.
Recent advances in 3D cell biology have enabled the reconstruction of
the intestinal stem cell niche in vitro. Since 2009, it has been
possible to maintain ISC in vitro in a long-term culture system known as
crypt culture or mini-gut culture (Pastula and Quante 2014; Sato et al. 2009).
Recently, such a mini-gut culture has been further improved by
incorporating the stromal microenvironment such as IMFs or neurons
(Lahar et al. 2011; Lei et al. 2014; Pastula et al. 2014, 2016a, 2016b) (Fig. 3).
For the stromal niche modeling in vitro, mesenchymal cells can be
either mixed together with epithelial cells and Matrigel (Pastula et al.
2016b) or epithelial organoids can be seeded on the mesenchymal cell monolayer (Holmberg et al. 2017; Lahar et al. 2011; Lei et al. 2014). In addition, IMFs and epithelial organoids can be seeded in separate layers in a Transwell (Pastula et al. 2016b).
Additionally, advances in 3D cell culture systems led to development of
intestinal organoid cultures derived from human embryonic stem cells
and human-induced pluripotent stem cells (Crespo et al. 2017; Rodansky et al. 2015), as well as intestinal organoids derived from large animal models (Khalil et al. 2016). Intriguingly, not only stromal cells, but also live bacteria such as Lactobacillus acidophilus (a part of the normal bacterial flora in our organism) can be added to the intestinal organoid cultures (Pierzchalska et al. 2017) (Fig. 3),
that provides an additional level of complexity to the epithelial
intestinal organoids, and offers a valuable tool to study interactions
between the gut microbiome and the intestinal epithelium. Since it is
possible to culture organoids derived from biopsy samples from patients
with colon cancer (van de Wetering et al. 2015) and IBD (Dotti et al. 2017),
such human-derived organoids could be used for the co-cultures with
different types of intestinal mesenchymal cells, immune cells and
microbiota, to better mimic organs for disease modeling in vitro.
Recently, a biobank of human-derived organoids derived from multiple
organs and also diseased tissue, including colon cancer and IBD, was
established (Dutta et al. 2017).
In addition, it would very useful to set up a living biobank of
different types of intestinal mesenchymal cells and gut microbiota
derived from patients suffering from colon cancer and IBD.
Certainly,
application of in vitro 3D organ models, such as those described above,
for further studies on the role of microenvironment–epithelial
interactions in the intestinal stem cell niche will lead in the future
to new exciting discoveries in both basic and translational research.
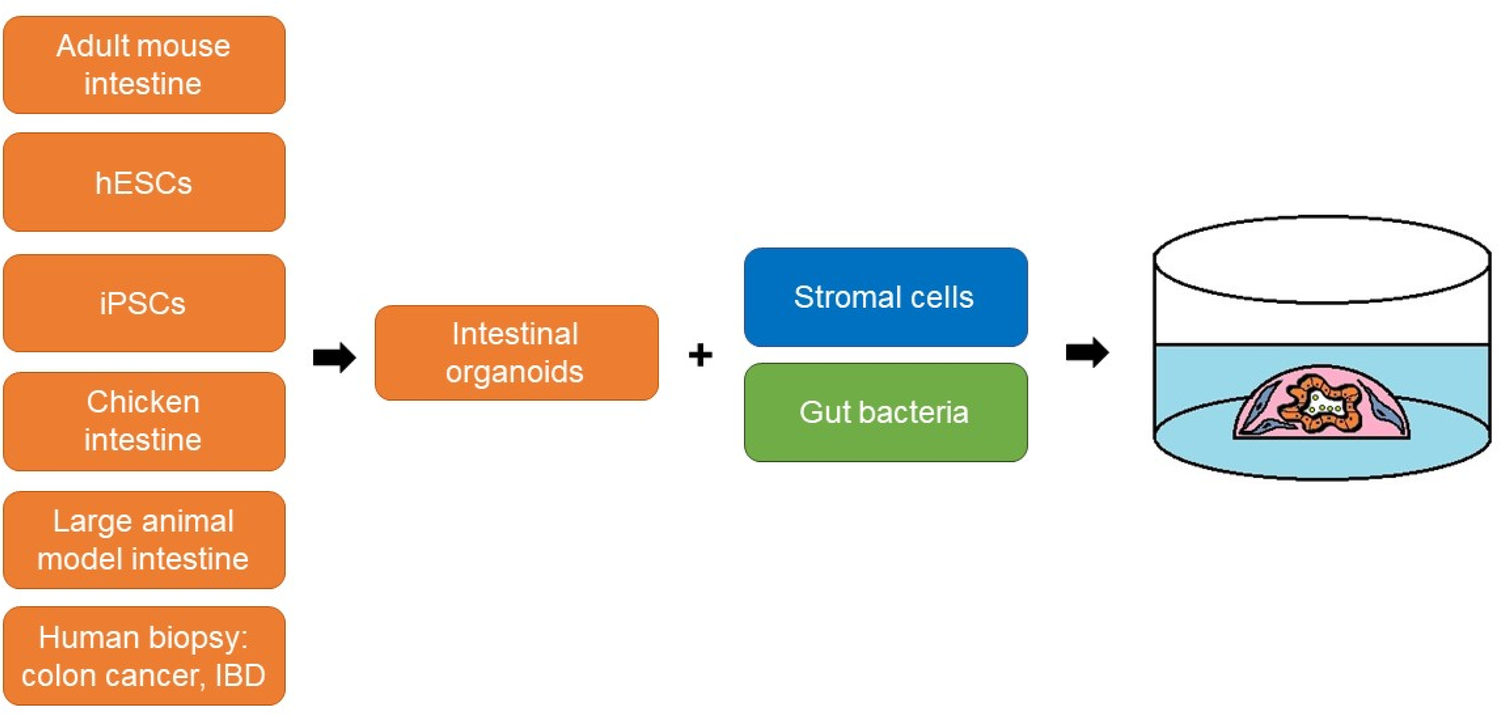
Fig. 3
Modifications
of the mini-gut culture system to reconstruct the intestinal tissue
microenvironment in vitro. For the co-culture, intestinal organoids can
be combined with stromal cells or/and live bacteria. A source of primary
intestinal epithelial cells can be, e.g., adult mouse intestinal
tissue, chicken intestinal tissue, human embryonic stem cells (hESCs),
induced pluripotent stem cells (iPSCs), and biopsy samples from patients
with colon cancer or inflammatory bowel disease (IBD)